This is a response to this creationist’s post on Human evolution, in which I found so many flaws that I have to at least correct the most obvious factual errors for general readers. At the same time, this may serve as a quick primer into neutral theory.
First, in every sperm or egg there are about 40 novel single nucleotide mutations (conservative estimate) that are passed from parent to offspring. We know this because we can sequence children and their parents and simply count the differences. So in the example in the post, with only one surviving couple each generation, the numbers of mutations passed on to the next generation is not 1 but 40.
If you now do the math (and still take the example in the post where only one couple survives each generation) and assume 500,000 generations since the common ancestor of chimps and humans, you get 20,000,000 substitutions on the human lineage, and 20,000,000 substitutions on the chimp lineage, amounting to a total of 40,000,000 differences between humans and chimps. That gives 1.3% sequence divergence and is very close to what has been reported in the Chimp Genome paper from 2005. So there is no dilemma!
But do we even need natural selection to explain the sequence divergence between humans and chimps? Let’s look at a scenario without natural selection, where all individuals in each generation survive and where there is no effect on fitness of any mutation whatsoever.
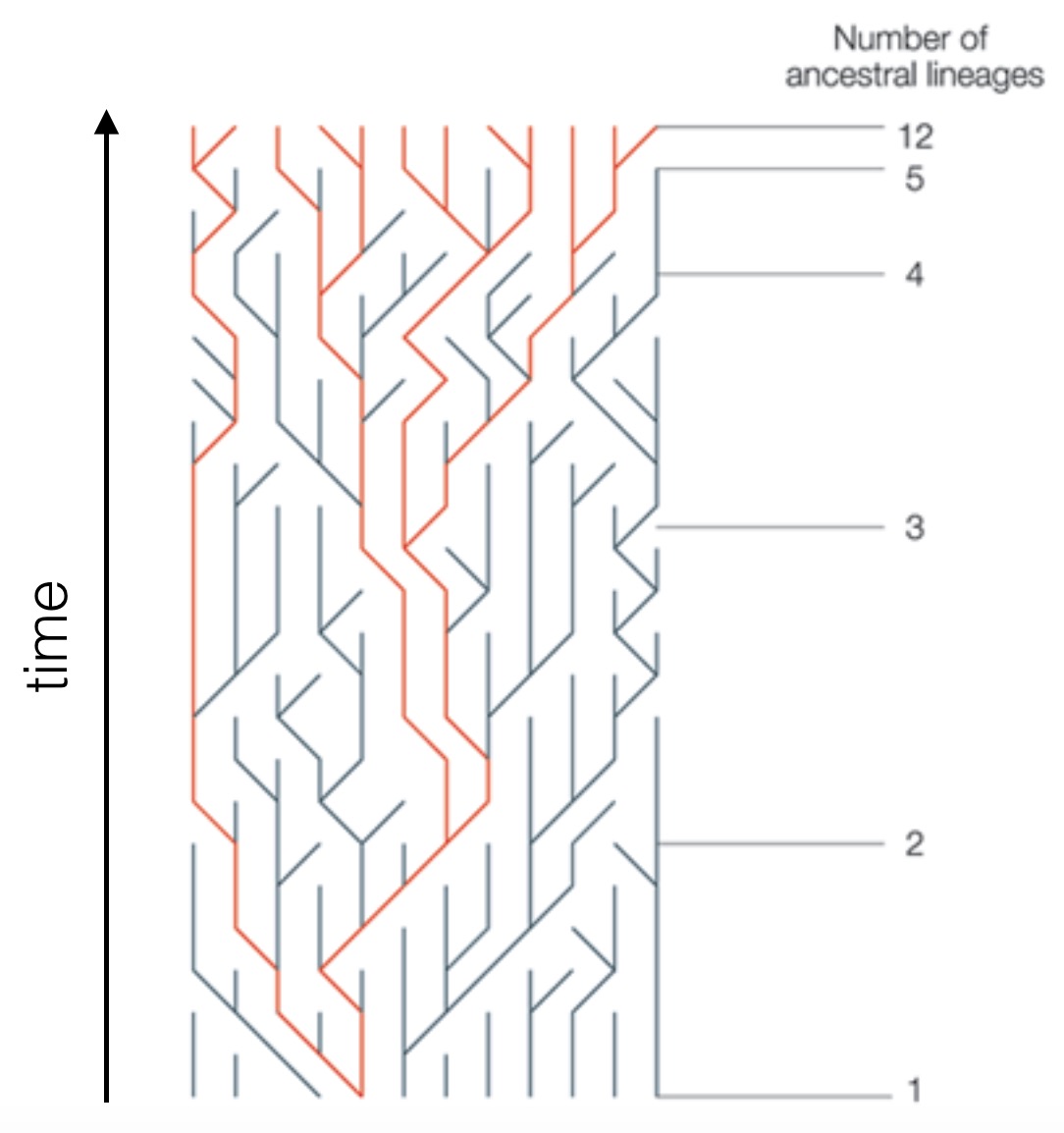
In this scenario, if there are N individuals, how many new mutations are there in the entire population each generation? Well, there should be 2N × 40 new mutations in total, because there are 2N sets of chromosomes in all diploid individuals (excluding X and Y chromosomes). What is the probability that any one of those mutations eventually becomes a substitution, so gets fixed in the population? For a neutral mutation, the answer is 1/2N, which can be seen from basic coalescence theory:
Every gene in the present generation has an equal chance of eventually becoming the sole ancestor for all genes many generations later, so every gene has a chance of 1/2N of being that ancestor. Let us look at how many mutations eventually get fixed each generation: we have 2N × 40 × 1/2N = 40.
So we find that in neutral evolution (where mutations have no fitness effects), the substitution rate equals the mutation rate. This remarkable result was derived by Motoo Kimura, who developed the neutral theory.
We find that even under strict neutrality, evolution progresses with a rate of 40 mutations per generation, so there is no need to invoke natural selection at all. Haldane’s paper from 1957 is therefore irrelevant if you want to explain human-chimp sequence divergence.
You should not confuse evolution with natural selection. Hardly any population geneticist believes today that a substantial fraction of substitutions between humans and chimps have been fixed by natural selection. In fact, we have to look pretty hard to find evidence for positive selection on the molecular level, because with few exceptions the phenotypic effects of mutations are typically small.
If you want to engage with science, you cannot just pick one paper from 1957 and ignore the large body of scientific work that came after it. Either you believe that science tells us something about the truth, or you don’t. If you do, you need to read and learn more than what is published in the Journal of Creation!